
![]()
Надотряд "Двоякодышащие" (Dipnoi, или Dipneustomorpha)
ОТРЯД РОГОЗУБООБРАЗНЫЕ (CERATODIFORMES)
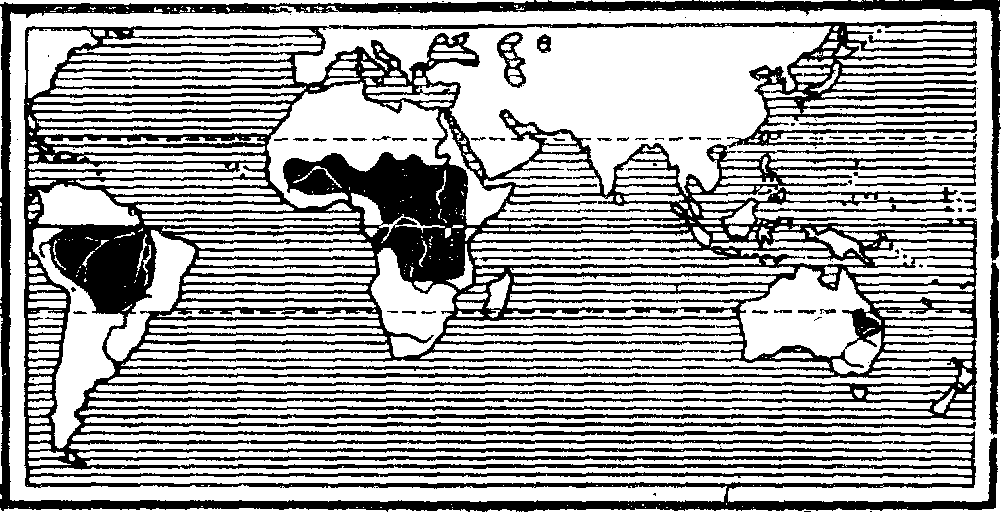
Рогозубообразные — единственная дожившая до нашего времени ветвь некогда многочисленных двоякодышащих рыб. Появившись в девонском периоде, двоякодьппащие рыбы процветали до триаса, после чего группа начала угасать. До нашего времени из двух отрядов двоякодышащиу. насчитывавших 11—12 семейств, сохранился только один отряд рогозубообразные с двумя семействами — рогозубовых (Ceratodidae) и чешуйчатниковых (Lepidosirenidae) — всего с шестью видами. Области распространения этих реликтовых форм —Южная Америка, тропическая Африка и Австралия — указывают на большую древность группы .
Современные двоякодытпащпе — типично пресноводные рыбы, прекрасно приспособленные к жизни в условиях пересыхающих и засушливый сезон водоемов.
Наиболее удивительно для двоякодышащих рыб так называемое “двойное” дыхание, откуда и произошло их название. Они способны осуществлять его благодаря тому, что помимо обычных для рыб жабр у них имеются еще и настоящие легкие, которые в существенных чертах своего строения сходны с легкими высших позвоночных. Эти легкие, заменяющие у них плавательный пузырь, соединяются с глоткой протоком, впадающим в нее с брюшной стороны. В связи с частичным переходом к легочному дыханию задненоздревые отверстия двоякодышащих рыб открываются в ротовую полость, образуя внутренние ноздри (хоаны), что позволяет им осуществлять дыхание атмосферным воздухом при закрытом рте; почти как у амфибий, имеется легочное кровообращение, т. е. венозная кровь поступает преимущественно в легкие, чему способствует также разделение предсердия неполной перегородкой. Теснейшим образом с легочным дыханием связано также наличие нижней полой вены. которая характерна для всех наземных позвоночных начиная с амфибий, но отсутствует у всех других рыб. кроме двоякодышащих.
Осевой скелет двоякодышащих- рыб во многом сохраняет примитивные особенности: тела позвонков отсутствуют, хрящевые основания верхних и нижних дуг сидят непосредственно на хорде хорошо сохраняющейся в течение всей жизни.Череп наряду с древними чертами характеризуется своеобразной специализацией. В хрящевой черепной коробке (нейрокранпум) развивается лишь одна пара замещающих костей (боковые затылочные). Имеется большое количество своеобразных покровных костей черепа. Небноквад;|атный -хрящ срастается с основанием черепа. На , сошнике, крылонебных костях и нижних челюгях сидят костные жевательные зубные пластин•;п. образующиеся от слияния многочисленных мелких зубов и весьма сходные с пластинками литно черепных (4 пластинки на верхней челюсти и 2 на нижней).
Хрящевой скелет парных плавников поддерживает почти всю лопасть плавника, кроме ее наружного края, где она поддерживается тонки•1И кожными лучами. Этот своеобразный внутрен•пш скелет состоит из длинной членистой центральчин оси, несущей у рогозубов (Ceratodidae) два ;1яда боковых членистых хрящевых элементов, | у чешуйчатников (сем. Lepidosirenidae) не имеющей этих придатков или несущей их рудименты. Внутренний скелет плавников соединяется с поясом лишь одним основным (базальным) члеником центральной оси и в этом отношении до известной тепени сходен с конечностью наземных позвоночных. Непарные плавники, спинной и анальный, полностью сливаются с хвостовым плавником. Последний симметричен, имеет дифицеркальное строение (у многих ископаемых двоякодыша|цих хвост был неравнолопастным — гетероцеркальным). Чешуя у древних форм была “космоидного” типа; у современных двоякодышащих верхний эмалевый слой и дентин утратились. В сердце имеется артериальный конус; кишечник снабжен спиральным клапаном,— это примитивные признаки. Мочеполовой аппарат сходен с таковым акуловых рыб и амфибий: имеется общее выводное отверстие (клоака).
Несмотря на то что по современным воззрениям двоякодышащие рыбы представляют собой боковую ветвь основного “ствола” водных позвоночных, интерес к этой удивительной группе животных не ослабевает, так как на ее примере можно проследить эволюционные попытки природы осуществить переход позвоночных животных от водного существования к наземному и от жаберного дыханин к легочному.
СЕМЕЙСТВО РОГОЗУБОВЫЕ, ИЛИ ОДНОЛЕГОЧНЫЕ (СКАТОШОАЕ)
К этому семейству относят несколько вымерших родов, ископаемые остатки которых находят на всех континентах, и близкий к ним современный род Neoceratodus, с одним видом. Характерны для них хрящевой нейрокраниум, наличие одного легкого и хорошо развитые пастообразные парные плавники, которые поддерживаются членистой центральной осью и двумя рядами боковых членистых лучей, отходящих от нее.
Единственный современный представитель семейства puzodi/v, или баррамун^а (Neoceratodus)
 встречается лишь в Квинсленде (Северо-Восточная Австралия), где он населяет бассейны рек Бёрнетт и Мэри. В последнее время он был пересажен также в некоторые озера и водохранилища Квинсленда, где и прижился. Рогозуб — крупная рыба,
встречается лишь в Квинсленде (Северо-Восточная Австралия), где он населяет бассейны рек Бёрнетт и Мэри. В последнее время он был пересажен также в некоторые озера и водохранилища Квинсленда, где и прижился. Рогозуб — крупная рыба,
Области распространения двоякодышащих рыб
достигающая в длину 175 см и массы свыше 10 кг. Его массивное тело сжато с боков и покрыто очень крупной чешуей, а мясистые парные плавники своими очертаниями чем-то напоминают ласты пингвинов. Окрашен в однообразные тона — от рыжевато-коричневого до голубовато-серого, которые несколько светлее на боках; брюхо обычно от беловато-серебристого до светло-желтого.
Рогозуб живет в реках с медленным течением и сильно заросших водной растительностью. Как и все рыбы, он дышит жабрами, но помимо этого каждые 40—50 мин поднимается к поверхности, чтобы подышать атмосферным воздухом. Выставив кончик рыла над водой, рогозуб с силой выбрасывает отработанный воздух при помощи единственного лёгкого,
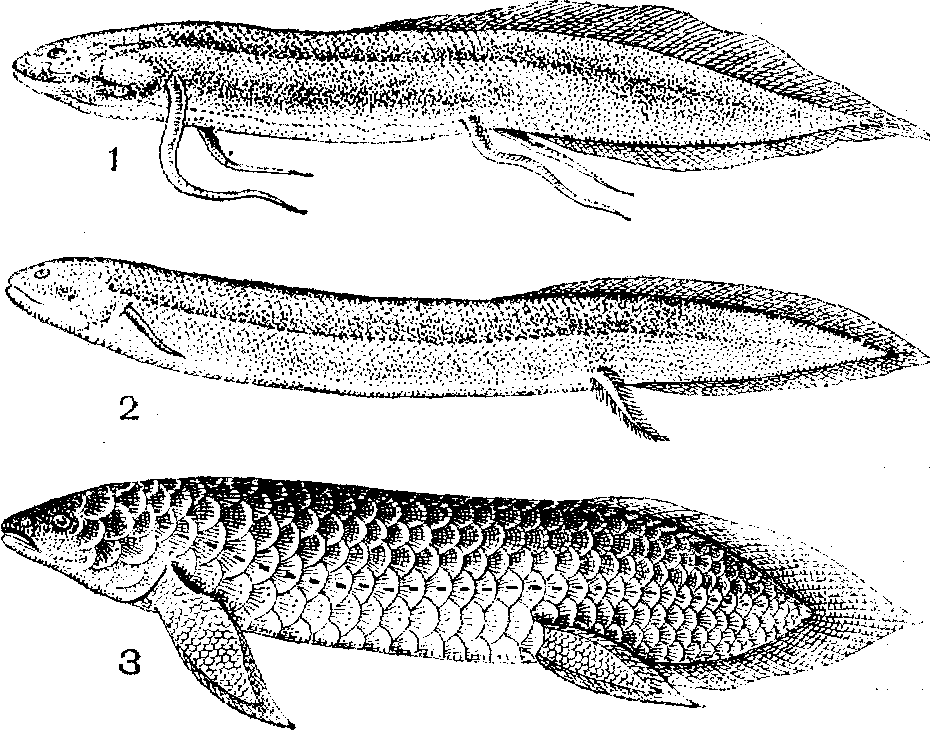
Двоякодышащие рыбы:
1 -— протоптер ; 2 — американский чешуйчатник;
3 — рогозуб
издавая при этом характерный стонуще-хрюкающий звук, который далеко разносится по окрестности. Тотчас вслед за этим, сделав глубокий вдох, он медленно погружается на дно. И выдох и вдох производятся им через ноздри при плотно сомкнутых челюстях. Нельзя не признать, что при дыхании атмосферным воздухом действия рогозуба напоминают действия китообразных. Даже находясь в воде, содержащей достаточное количество кислорода, рогозуб, по-видимому, не может довольствоваться жаберным дыханием и дополняет его дыханием легочным. Последнее особенно полезно для него в засушливые сезоны, когда русла рек полностью пересыхают на больших пространствах и когда вода сохраняется лишь в наиболее глубоких ямах (бочагах). В таких постепенно усыхающих убежищах, ища спасения, скапливается множество рыб, и в том числе рогозубы. Когда в перегретой стоячей воде в результате гнилостных процессов исчезает почти весь кислород и все остальные рыбы дохнут от удушья, рогозуб продолжает благоденствовать, перейдя на дыхание атмосферным воздухом. И даже тогда, когда при затянувшейся засухе эти убежища превращаются в кладбище для всего живого, а вода в них — в зловонную жижу, в которой разлагаются сотни трупов погибших животных,— даже и тогда рогозуб выживает, дождавшись спасительных дождей. Однако полное высыхание водоема гибельно и для него, так как он не может впадать в спячку, зарывшись в грунт, подобно своим африканским и южноамериканскому сородичам.
Вытащенный из воды рогозуб совершенно беспомощен и погибает скорее, чем многие другие рыбы, лишенные легких. Рогозуб — вялое и малоподвижное животное.Большую часть времени он обычно проводит на дне глубоких омутов, где лежит на брюхе или стоит, опираясь на парные плавники и на хвостовую часть тела. В поисках пищи он медленно ползает на брюхе, а подчас и ходит, опираясь на те же парные плавники. В толще воды он, как правило, медленно передвигается за счет едва заметных изгибаний своего тела. Только если его вспугнуть, рогозуб пускает в ход свой мощный хвост и выказывает свою способность к быстрому движению. Судя по всему, суточный ритм у этого животного выражен слабо и нередко рогозуб проявляет свою вялую активность в любое время дня и ночи. Его пищу составляют различные беспозвоночные (моллюски, ракообразные, личинки насекомых, черви и т. п.). Правда, кишечник рогозуба обычно набит мелко пережеванными растительными остатками, но, по всей видимости, растительная пища им не усваивается, а захватывается вместе с беспозвоночными животными. По крайней мере в неволе он безо всякого ущерба довольствуется “скоромной” пищей, не выказывая потребности в “вегетарианской” диете.
Нерест рогозуба сильно растянут и продолжается с апреля по ноябрь. Наиболее интенсивно он идет в сентябре —октябре, когда наступает период дождей, реки вздуваются и вода в них хорошо аэрируется. Рогозуб откладывает икру на водную растительность и не проявляет дальнейшей заботы о потомстве. Так как оболочка у икринок не клейкая, то многие из них скатываются и падают на дно; не совсем ясно, как это сказывается на их выживании. Икринки довольно крупные, они достигают в диаметре 6,5—7,0 мм и заключены в студенистую оболочку, что делает их весьма сходными с лягушачьей икрой. Это сходство усугубляется большим количеством желтка и особенностями эмбрионального развития. Развитие икринок продолжается 10—12 суток. В отличие от личинок чешуйчатников и протоптеров, у личинок рогозуба совершенно отсутствуют наружные жабры и цементный орган. До того как у них рассосется желточный мешок, они неподвижно лежат на боку на дне и лишь время от времени, как бы встрепенувшись, перескакивают на другое место поблизости, чтобы снова замереть в прежнем положении. С переходом к активному питанию личинки держатся тихих и мелких заводей, где поначалу питаются нитчатыми водорослями, переходя со временем на питание беспозвоночными. Грудные плавники появляются у них, как правило, на 14-й день после выклева, а брюшные — много позже (примерно через два с половиной месяца).
Рогозуба употребляют в пищу, и красноватое мясо его очень ценят как аборигены, так и белые поселенцы. Рогозуб хорошо ловится на крючок в любое время суток, но бывают периоды, длящиеся до недели и более, когда он не берет ни на какую наживку. Очень искусно ловят (вернее, ловили) рогозуба аборигены, которые используют для этой цели небольшие самодельные сетки. Взяв в каждую руку по такой сетке, рыболов ныряет в глубокую яму, стремясь обнаружить рыбу, лежащую на дне. Осторожно подведя сетки одновременно к голове и хвосту рогозуба, рыболов захватывает ими рыбу и всплывает с ней на поверхность. Вряд ли какая-либо другая рыба проявляет такую инертность, чтобы дать захватить себя голыми руками. Даже прикосновение не всегда вспугивает рогозуба. А если его все же побеспокоить, то и тогда, все еще не ощущая опасности, он пускает в ход свой сильный хвост и резким рывком уходит от назойливого рыбака, чтобы снова неподвижно залечь неподалеку. В таком случае возобновить преследование ничего не стоит. По-видимому, такое пренебрежение к опасности выработалось у рогозуба в то время и в тех условиях, когда у него не было врагов и ему некого было опасаться. Лишь попав в тенета или на крючок, флегматичный рогозуб проявляет недюжинную силу и яростно борется за свою жизнь. Но на долгое сопротивление он не способен: его неистовство быстро истощается, и он безвольно отдается на волю победителя.
В неволе это мирное животное хорошо уживается с другими рыбами и с себе подобными. С рогозубом связана одна из самых удивительных мистификаций, какие только знает зоология. Начало ее восходит к августу 1872 г. В это время директор Брисбейнского музея совершал поездку по северному Квинсленду. Однажды ему сообщили, что в его честь приготовили завтрак и что ради него туземцы не поленились доставить к столу очень редкую рыбу, пойманную ими в 8—10 милях от того места, где должно было произойти пиршество. Польщенный директор принял это предложение и действительно увидел рыбу очень странного облика: ее длинное массивное тело было покрыто мощной чешуей, плавники походили на ласты, а рыло напоминало утиный клюв. Прежде чем воздать должное столь необычному блюду (нет нужчы говорить, что рыба была уже приготовлена), директор сделал с него набросок, а вернувшись в Брисбейн, передал его Ф. де Кастельнау, тогдашнему ведущему австралийскому ихтиологу. Кастельнау не замедлил по этому рисунку описать новый род и вид Ompax spatuloides, которого он отнес к двоякодышащим рыбам. Эта публикация вызвала довольно бурную дискуссию о родственных связях Ompax и о его месте в классификационной системе. Оснований для споров было много, так как в описании Опарах многое оставаясь неясным и совсем отсутствовали сведения по анатомии.
Попытки добыть новый экземпляр оказались безрезультатными. Как и всегда, нашлись скептики, которые высказывали сомнение в существовании этого животного.
Тем не менее таинственный Ompax spatuloides •')ез малого в течение 60 лет продолжал упоминаться во всех справочниках и сводках по австралийской фауне.
Загадка разрешилась неожиданно. В 1930 г. в < Сиднейском бюллетене” появилась заметка, автор которой пожелал остаться неизвестным. В этой
заметке сообщалось, что с простодушным директором Брисбейнского музея сыграли невинную шутку, так как поданный ему “Ompax” был приготовлен из хвостовой части угря, туловища кефали, головы и грудных плавников рогозуба и рыла утконоса. Сверху все это хитроумное гастрономическое сооружение было искусно покрыто чешуей того же рогозуба.
Так Ompax spatuloides был вычеркнут из фаутшстических списков, л рогозуб остался единстленным ныне живущим преде га вителом двоякодышащих в Австралии.
СЕМЕЙСТВО ЧЕШУЙЧАТНИКОВЫЕ, ИЛИ ДВУЛЕГОЧНИКОВЫЕ (LEPIDOSIRENIDAE)
Чешуйчатниковые характеризуются удлиненным угреобразным телом, которое вплоть до брюшных плавников округло в поперечном сечении. Они имеют парное легкое; мелкая циклоидная чешуя, покрывающая их тело и отчасти голову, глубоко спрятана под кожу, а их гибкие парные плавники имеют жгутовидную форму. Наиболее характерна для рыб этого семейства способность в течение всей своей жизни существовать во временных водоемах, нередко полностью пересыхающих в засушливый сезон, дл-ящийся подчас до 9 месяцев. На все это время они впадают в спячку, зарывшись в грунт и полностью переходя на дыхание атмосферным воздухом. В этом семействе насчитывается 5 видов: 4 вида, обитающих в тропической Африке, принадлежат к роду протоптеров (Protopterus), а южноамериканский род чешуйчатников (Lepidosiren) представлен всего одним видом. Близость между южноамериканским и африканским представителями пресноводных двоякодышащих рыб является веским доводом в пользу существования сухопутной связи между Африкой и Южной Америкой в далеком прошлом.
Пожалуй, самое существенное отличие протоптеров от чешуйчатников сводится к тому, что у первых имеется 6 жаберных дуг и 5 жаберных ще- лей, в то время как у вторых — всего 5 жаберных дуг и 4 жаберные щели. Иногда их рассматривают как представителей особых семейств (Lepidosirenidae и Protopteridae).
Четыре вида рода протоптеры (Protopterus) внешне очень сходны и отличаются друг от друга по своей окраске, по количеству ребер, по степени развития и ширине кожной оторочки парных плавников и другим признакам.
Наиболее крупный вид — большой протоптер (Protopterus aethiopicus, местное название “мамба”) — достигает в длину свыше 2 м, окрашен в голубовато-серые тона. с многочисленными мелкими темными пятнами, иногда образующими “мраморный” узор (рис. 48). Этот вид обитает от Восточного Судана до озера Танганьика.
Малый протоптер (Р. amphibius), по-видимому,самый мелкий вид, не превышающий в длину 30 см. Обитает в дельте Замбези и в реках к юго-востоку от озера Рудольфа. Для молоди этого вида характерно наличие трех пар наружных жабр, которые сохраняются в течение очень длительного времени.
Темный протоптер (Р. do! lot), обитающий только в бассейне Конго, характеризуется наиболее удлиненным телом и очень темной окраской. Достигает в длину 85 см. Внешне этот вид в наибольшей степени сходен с южноамериканским чешуйчатником.
Бурый протоптер (Р. annectens), достигающий в длину 90 см,—обычная двоякодышащая рыба Западной Африки. Он населяет бассейны рек Сенегала, Гамбии, Нигера и Замбези, озеро Чад и район Катанги. Спина у этого вида обычно бурозеленая, бока светлее, брюхо грязно-белое. Наиболее хорошо изучена биология именно этого вида. Для климата тропической Африки характерна резкая смена дождливого и засушливого сезонов. Дождливый сезон начинается в мае—июле и длится 2—3 месяца, а всю ос1альную часть года длится засушливый период. В период бурных тропических лнкнрй реки взбухают и разливаются, заэопляя огромные площади низин, в которых вода держится 3—5 месяцев в юду. В эти временные водоемы, где в изобилии имеется легко доступная пища, из рек устремляется масса рыб, но по мере их усыхания, спасаясь от гибели, рыбы возвращаются в реки до того, как обмелеют протоки. Совершенно иначе ведет себя протоптер. Оказывается,что в реках он, как правило, вообще не обитает, а постоянно живет в таких временных водоемах и весь его жизненный ритм теснейшим образом связан с их гидрологическими особенностями.
Местные рыбаки бассейна реки Гамбии, хорошо знающие повадки протоптера, недаром говорят: “Камбона (так они называют протоптера) — необыкновенная рыба: она не уходит вслед за водой, а вода сама приходит к ней”. В дождливое время протоптер ведёт в водоемах активный образ жизни — питается, размножается и растет. А в засушливый период впадает в спячку, проводя ее в специально устроенных гнездах.
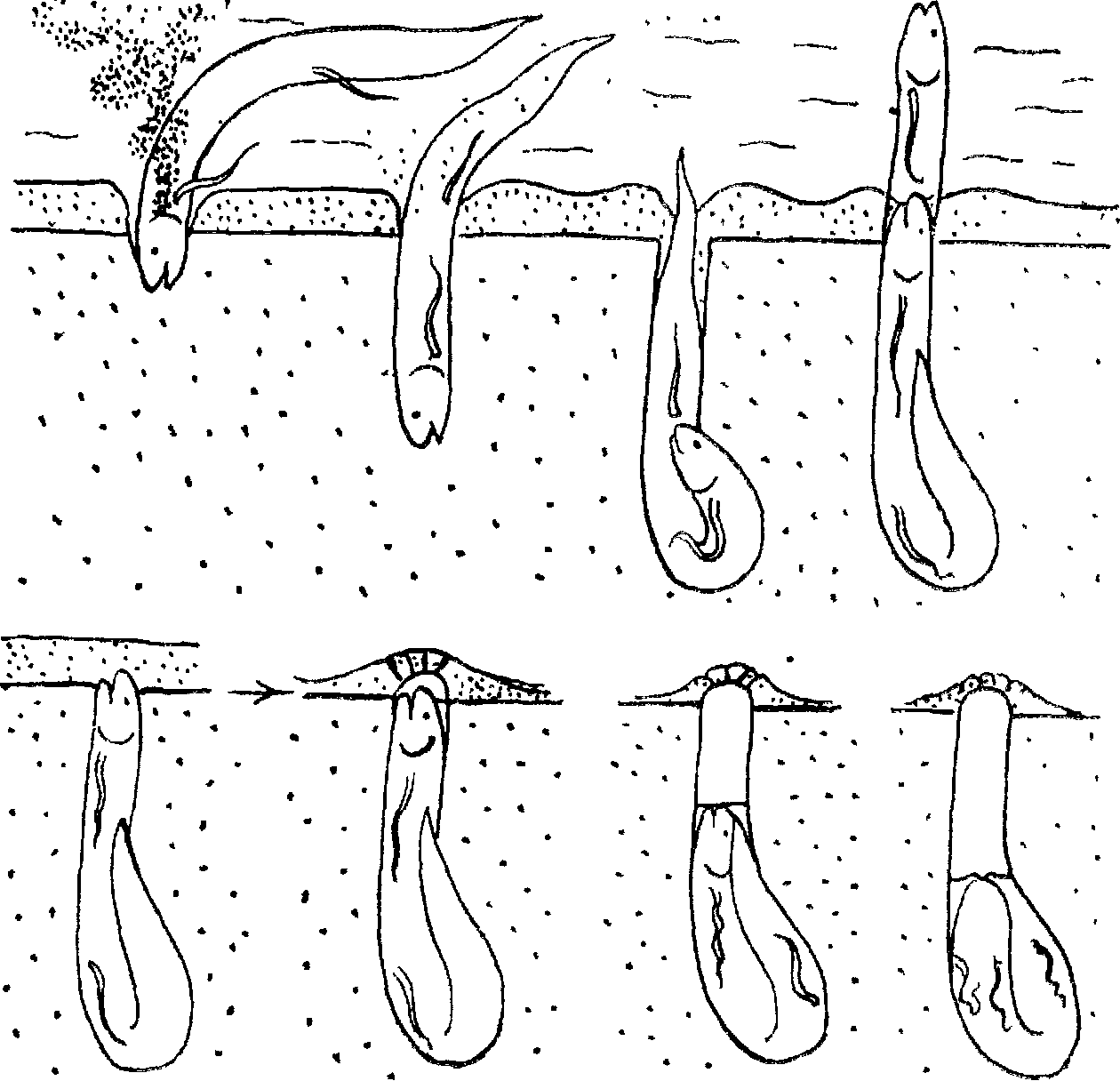
С наступлением засушливого сезона и по мере высыхания временных водоемов протоптеры начинают готовиться к спячке: крупные рыбы делают это, когда уровень воды понижается до 10 см. а более мелкие — когда слой воды не превышает 3—5 см. Обычно в таких водоемах дно покрыто мягким плом, в котором содержится большое количество растительных остатков. Под слоем ила, достигающим толщины 2,5—5 см, лежит плотная глина с примесью мелкого песка. Свое “спальное гнездо” протоптер роет ртом. Засосав в ротовую полость очередную порцию ила , он с силой выбрасывает его вместе с водой через жаберные отверстия. Мягкий ил легко поддается “бурению”, но подлежащий слой плогной глины рыть намного трудней. Совершая всем телом энергичные плавательные движения, рыба упирается рылом в грунт и выгрызает кусочек глины. Откушенный кусочек пережевывается, выбрасывается с водой через те же жаберные отверстия и выводится из норы в виде облачка мути с восходящими юками воды, создаваемыми за счет изгибаний jp,га. Благодаря этому более крупные частицы измельченной глины оседают в непосредственной близости от входною отверстия, что имеет cyщественное значение для создания завершающего постройку предохранительно]о колпачка. Дойдя до нужной глубины, рыба расширяет нижнюю часть норы (“спальню”) ровно настолько, чтобы иметь возможность, сложившись пополам, перевернуться в ней головой вверх. Теперь “спальное гнездо” почти готово, и животное ждет полного спада воды, выставив рыло из входного отверс1ия и время от времени приподнимаясь к поверхности для дыхания атмосферным воздухом. Когда пленка усыхающей воды доходит до верхней кромки жидкого ила, выстилающего дно водоема, тогда благодаря дыхательным движениям, производимым рыбой, часть выброшенной у входного отверстия глины засасывался в нею и закупоривает выход. После этою животное уже не выходит на поверхность. До того как эта “пробка” окончательно просохнет, протоптер, тычась в нее рылом, уплотняет ее снизу и несколько приподнимает в виде колпачка, нередко имеющего трещины. Колпачок маскирует “спальное гнездо” и предохраняет его от засорения, будучи достаточно прочным, чтобы противостоять разрушению. Одновременно с этим примесь из мелких песчинок делает его достаточно пористым, чтобы пропускать воздух, чему еще больше способствуют трещины. Как только колпачок затвердевает, вода н норе становится вязкой от обилия слизи, выделяемой протоптером. По мере просыхания грунта уровень воды во входной камере постепенно падает, в результате чего она превращается в воздушную камеру, а рыба, послушно следуя за зеркалом воды, все ниже и ниже опускается в расширенную нижнюю часть норы, т. е. в “спальню”, где, наконец, и замирает в характерном для нее положении.
Удивительное чувство испытывает приезжий натуралист. когда в сопровождении местных жителей он впервые отправляется на поиски “спальных гнезд” протоптера. Трудно поверить, что потрескавшаяся от зноя равнина, покрытая выжженной растительностью, еще недавно была дном водоема и что где-то рядом в окаменевшей земле спят сотни и тысячи рыб. Он бывает сильно удивлен, когда туземцы, чуть не ползая на коленях, начинают тщательно, дюйм за дюймом обследовать почву. Вскоре выясняется, что они ищут небольшие возвышеньица диаметром 5—15 см, которые чт окружающей их почвы, окрашенной в более и. in менее серые тона, отличаются коричневатым оттенком. Достаточно одного удара мотыгой, что-бы под таким срезанным бугорком обнаружилась идущая вглубь нора. Иными словами, каждый такой холмик представляет собой так называемую предохранительную крышку, или колпачок, коюрый прикрывает сверху вход в “спальное гнездо” протоптера. Опытным глазом эти холмики можно обнаруживать без труда. Лишь у мелких рыб, длиной менее 15 см, они столь слабо выражены, что их практически невозможно найти.
Круглый ход, обычно идущий вертикально вниз, имеет гладкие стенки. Это так называемая воз1ушная камера. Ее диаметр колеблется от 5 до 70 мм, а длина — от 30 до 250 мм. Эти размеры занпсят только от величины рыбы, залегшей в спячку. Даже длина воздушной камеры не зависит от гого, на глубоком или на мелком месте строилось • гнездо”. Внизу воздушная камера постепенно расширяется и переходит в так называемую “спальчю”, где и покоится заключенная в кокон рыба. V крупных рыб “спальня” залегает на глубине до полуметра.
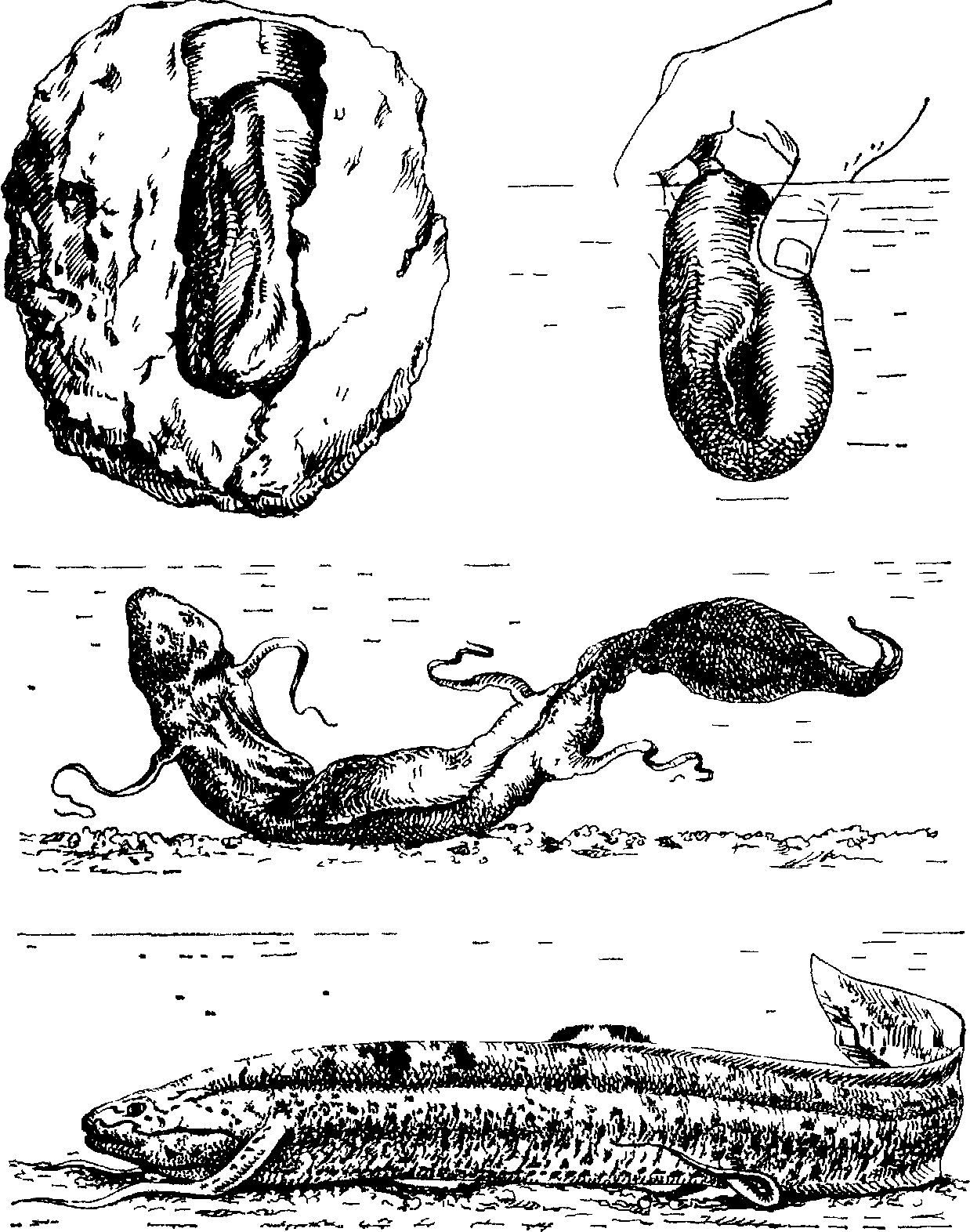
 Спящий протоптер, как правило, принимает строго определенное положение. Его рыло всегда направлено вверх, а тело складывается пополам так, что изгиб приходится посредине между груд-
Спящий протоптер, как правило, принимает строго определенное положение. Его рыло всегда направлено вверх, а тело складывается пополам так, что изгиб приходится посредине между груд-
ными и брюшными плавниками, иначе говоря, эти плавники оказываются рядом и на одном уровне.
Сложенные передняя и задняя части тела очень тесно прижаты друг к другу, а уплощенный хвост
перехлестнут через верхнюю часть
Зарыванпе протоптера в грунт
головы и столь же плотно прижат к спине. При этом нижняя кромка хвоста, полностью прикрывающего глаза, проходит вдоль края верхней челюсти, оставляя свободным слегка приоткрытый рот. Свернувшаяся таким образом рыба заключена в своеобразный кокон. В мире рыб это уникальное образование способны создавать только представители рода Protopterus.
Кокон представляет собой тончайшую пленку толщиной 0,05—0,06 мм, образующуюся при затвердевании слизи, которую выделяет приготовившаяся к спячке рыба. Его стенки состоят из муцина с небольшой примесью неорганических соединений (их основу составляют карбонат и фосфат кальция), перешедших из почвы в момент образования кокона. Кокон — цельное образование (без каких-либо перетяжек) и столь плотно обтягивает спящего протоптера, что между его стенками и телом рыбы не остается никаких зазоров. Сморщенные парные плавники спящей рыбы очень сильно вжимаются в тело и не оставляют никаких следов на внутренней стенке кокона. Округлая верхняя часть кокона, повторяющая контур стенок воздушной .камеры в месте перехода ее в “спальню”, уплощена и слегка всхолмлена непосредственно над ртом рыбы. Это поднятие имеет небольшое углубление на вершине, в центре которого
находится отверстие воронкообразной, трубочки длиной 1—5 мм, ведущей прямо в приоткрытый рот спящего протоптера. Как раз через это крошечное дыхательное отверстие и осуществляется единственная связь рыбы с наружной средой. Обычно кокон окрашен под цвет рыжевато-бурого грунта за счет красящих неорганических веществ, содержащихся в почве. В тех случаях, когда эти вещества отсутствуют, кокон может быть прозрачным, как целлофан. Его внутренняя стенка всегда влажная, так как тело рыбы остается покрытым слизью до окончания спячки.
Способность протоптера “облачаться” в кокон на время спячки столь необычна и удивительна, что первые исследователи, увидевшие этот кокон. никак не могли поверить своим собственным глазам. Вопреки явной очевидности они принимали стенки кокона за засохшие листья, предположив, что отходящая ко сну рыба заворачивается в них, наклеивая их на себя с помощью густой слизи.
Так, завернутым в фантастические листья, как в некое подобие пеленок, и был изображен спящий протоптер в публикации Джердайна, появившейся в 1841 г. И это не было шуткой. Вполне естественно, что для поддержания своей жизнедеятельности спящий в коконе протоптер должен не только дышать, потребляя кислород, но и питаться, т. е. расходовать какие-то запасы “топлива”, и что-то делать с продуктами распада, избыток которых в организме обычно приводит к смертельному исходу.
В противоположность всем другим позвоночным животным, впадающим в спячку, протоптер, заключенный в кокон, расходует не жировые запасы, а свои собственные мышечные ткани. В начале спячки обмен веществ происходит у него еще на довольно высоком энергетическом уровне, но
постепенно он замирает и протекает в дальнейшем в очень экономном режиме, так как, в противном случае, ему не хватило бы “топлива”, т. е. мышечных тканей. За время спячки протоптер сильно теряет в массе. Так, например, рыба длиной 40 см, массой 374 г после шестимесячного пребывания в коконе имела длину 36 см и массу 289 г, т. е. потеряла в массе более 20% и уменьшилась в размерах на 10%. Такие сравнительно большие потери объясняются тем, что во время спячки ткани протоптера расходуются не только на поддержание жизнедеятельности организма, но и на созревание гонад. Потери восполняются довольно быстро: та же рыба уже через месяц восстановила свою массу и достигла прежних размеров.
Во время спячки протоптера вся вода, образующаяся при расщеплении белков, теряется при дыхании и моча не выделяется (да ее и некуда было бы вывести, так как рыба заключена в кокон, плотно облегающий ее тело). Поэтому образующаяся мочевина в огромных количествах скапливается в организме, составляя к концу спячки 1—2% массы тела, что следует рассматривать как удивительный физиологический парадокс: на большинство позвоночных животных избыток мочевины в организме действует как сильнейший яд, и смерть наступает при ее концентрации, в 2 тыс. раз меньшей, чем у спящего протоптера, которому она не причиняет никакого вреда. Уже через несколько часов после выхода протоптера в воду весь избыток мочевины выводится из организма через жабры и почки.
В зависимости от местных условий, значительно колеблющихся в разные годы, протоптер проводит в спячке 6—9 месяцев. Любопытный рекорд был побит бурым протоптером, который в экспериментальных условиях провел в беспрерывной спячке свыше четырех лет без каких-либо вредных последствий для себя. Однако в тех случаях, когда водоемы не пересыхают, протоптеры в спячку не впадают. Этого легко добиться и в аквариумных условиях. Тем не менее было замечено, что “бодрствующие” в течение ряда лет протоптеры, содержащиеся в аквариуме (один из них провел без спячки 13 лет), время от времени становятся вялыми, малоподвижными и даже отказываются от пищи. Такое состояние замечается у них в среднем один раз в году и продолжается от нескольких недель до 2—3 месяцев без каких-либо признаков заболевания.
Почти несомненно, что такое поведение связано с прирожденной привычкой впадать в спячку и что спячка составляет неотъемлемую часть жизненного ритма этих рыб. Ради точности следует добавить, что эти наблюдения производились над особями бурого протоптера, пойманными в бассейне Гамбии, где этот вид обычно впадает в спячку. Возможно, что у протоптеров других видов такая ритмика выражена не столь резко. Известно, например, что в Великих озерах Центральной Африки протоптеры в ежегодную спячку не впадают, так как для этого у них нет необходимости и соответствующих условий.
С наступлением периода дождей пересохшие водоемы быстро наполняются водой и протоптеры возвращаются к активной жизни из своего добровольного заточения. Самый процесс их пробуждения в природе пока еще не прослежен, но о нем можно судить по специальному эксперименту, поставленному в 1931 г. (рис. 5). Этот простой опыт заключался в том, что вырубленные из грунта куски глины с заключенными в них протоптерами зарывались в мелкую лужу так, чтобы слой воды над ними не превышал 5 см. Приблизительно через час у выходного отверстия показалась первая рыба. После короткой рекогносцировки она поднялась к поверхности воды и жадно заглотнула воздух, чтобы тотчас после эюго скрыться в гнезде. Поначалу эти действия повторялись через каждые 3—5 мин^ но постепенно интервалы между очередными выходами к поверхности удлинились до обычных 10—20 мин. При этом рыба все меньше и меньше пряталась в гнезде, пока через 6—7 ч совем не покинула его.
Замечено, что, чем дольше длится спячка у про•штера, тем больше времени требуется ему для того, чтобы стряхнуть с себя сон. В течение первых нескольких дней рыбы, проведшие в спячке 6—8 месяцев, плохо контролируют свои движения, передвигаясь резкими и неуклюжими рывками, подобно калекам. При этом хвост у них доъольно продолжительное время остается отогнутым вверх и несколько вбок, а смятые парные плавники лишь постепенно распрямляются и приобретают упругость.
Протоптер — всеядная рыба. Основу его пищи составляют разнообразные моллюски, крабы, креветки и отчасти рыбы. Захватив добычу, он не глотает ее, а выбрасывает изо рта, придерживая за самый кончик, и начинает методично пережевывать, пока она вся не скроется во рту. Затем снова выплевывает ее и снова жует. И так несколько раз. Добычу он не хватает, а засасывает, причем делает это с непостижимой быстротой и проворством. Возможно, что как раз при этом захватываются и отдельные части растений, остатки которых нередко находят у него в желудке.
На тех, кто наблюдал протоптеров в аквариуме эти рыбы производят впечатление вялых и малоподвижных животных. Но это впечатление обманчиво, так как протоптеры ведут ночной образ жизни и выходят на охоту с наступлением темноты. В это время их активность резко возрастает, и они чаще поднимаются к поверхности для дыхания атмосферным воздухом. Перемещаются протоптеры двояким способом: они или плавают за счет угреобразного изгибания тела, или же передвигаются по дну и среди донной растительности с помощью парных плавников, причем, помимо двигательных функций, эти плавники играют большую роль при отыскании добычи, так как они густо усеяны вкусовыми почками (особенно обильно покрыты ими грудные плавники). Стоит себе представить протоптера, охотящегося ночью среди
густых зарослей водной растительности в мутной воде, чтобы понять, какую незначительную роль может играть в этих условиях зрение. Здесь и приходят на помощь длинные и гибкие парные плавники, которыми ползущая рыба обследует “на вкус” окружающее ее пространство. Стоит протоптеру лишь коснуться съедобного объекта одним из четырех плавников, как молниеносным броском он подскакивает к добыче и отправляет ее в рот.
 Развитие гонад у протоптера начинается тотчас же после нереста, причем большая часть времени при их созревание приходтся на период спячки. Уже в августе — сентябре, i. с. inyciH месяц полтора после начала сезона дождей и окончания спячки, начинается нерест, который длится около месяца. К этому времени сооружается специальное выводковое гнездо. Оно строится обычно на мелководье, где слой воды не превышает 40—50см и где дно поросло густой травой, нередко достигающей в высоту 2 м. Как правило, такое гнездо представляет собой подковообразную нору с двумя входными отверстиями. Одно из них — более широкое — имеет в диаметре 20—30 см, а другое, более узкое,— только 10—15 см. В нижней части этой норы, лежащей примерно на глубине 40 см от поверхности грунта и наиболее удаленной от входных отверстий, находится расширенная выводковая камера, в которой откладывается икра и держатся личинки. Иногда гнезда имеют по три входных отверстия, ведущих в общую выводковую камеру, или всего лишь один выход, когда для постройки гнезда используются крутые кочки или искусственные земляные насыпи, разделяющие рисовые поля. Стенки гнезда слизью не покрываются и ничем специально не укрепляются: от обвала его предохраняет плотный грунт, скрепленный многочисленными корнями растений. Никакой подстилки в выводковой камере нет, и икра откладывается непосредственно на сеглинистое дни Т -f- ьак гнезда строятся на мелководье то, :п'-
3
'
-
-i-г^бы добираться до более глубокой вон- т птеры проделывают своеобразные -: ... приминая и раздвигая густую траву. "^ -- по этим “тропам” и находят выводковые
Развитие гонад у протоптера начинается тотчас же после нереста, причем большая часть времени при их созревание приходтся на период спячки. Уже в августе — сентябре, i. с. inyciH месяц полтора после начала сезона дождей и окончания спячки, начинается нерест, который длится около месяца. К этому времени сооружается специальное выводковое гнездо. Оно строится обычно на мелководье, где слой воды не превышает 40—50см и где дно поросло густой травой, нередко достигающей в высоту 2 м. Как правило, такое гнездо представляет собой подковообразную нору с двумя входными отверстиями. Одно из них — более широкое — имеет в диаметре 20—30 см, а другое, более узкое,— только 10—15 см. В нижней части этой норы, лежащей примерно на глубине 40 см от поверхности грунта и наиболее удаленной от входных отверстий, находится расширенная выводковая камера, в которой откладывается икра и держатся личинки. Иногда гнезда имеют по три входных отверстия, ведущих в общую выводковую камеру, или всего лишь один выход, когда для постройки гнезда используются крутые кочки или искусственные земляные насыпи, разделяющие рисовые поля. Стенки гнезда слизью не покрываются и ничем специально не укрепляются: от обвала его предохраняет плотный грунт, скрепленный многочисленными корнями растений. Никакой подстилки в выводковой камере нет, и икра откладывается непосредственно на сеглинистое дни Т -f- ьак гнезда строятся на мелководье то, :п'-
3
'
-
-i-г^бы добираться до более глубокой вон- т птеры проделывают своеобразные -: ... приминая и раздвигая густую траву. "^ -- по этим “тропам” и находят выводковые
-:- .л. так как в мутной воде среди буйной рас-t'точности обнаружить входное отверстие каким-либо иным способом очень трудно, если только случайно не провалиться в него. Нередко “тропы” тянутся на несколько метров, а когда уровень воды резко понижается (что случается довольно часто), то протоптерам приходится добираться до воды по суше. Но даже при очень резких колебаниях уровня воды сами гнезда никогда не обсыхают. Местами такие гнезда располагаются в недалеком соседстве друг от друга на расстоянии 7-8 м.
Всю заботу об охране гнезда и потомства берет на себя самец. Он самоотверженно защищает свое гнездо и злобно кусает всякого, кто посмеет приблизиться к нему, не отступая и перед человеком (его яростных атак туземцы боятся). Если даже его выгнать из гнезда палкой, то он бесстрашно возвращается через несколько минут. Спрятавшись в одном из отнорков, самец поддерживает постоянный ток воды в выводковой камере за счет ундулирующих движений хвоста. Свою заботу о потомстве он прекращает лишь тогда, когда личинки покидают гнездо.
Никому не довелось наблюдать процесс постройки гнезда, и до сих пор неизвестно, строит ли его самец или самка, или же они строят его совместно. Судя по тому, что в охране гнезда и потомства самка никакого участия не принимает, предпочтительней думать, что гнездо строит самец. Икринки протоптера имеют в диаметре 3,5—4,0 мм. Их количество в одной кладке достигает 5 тыс., по нередки случаи, когда их бывает значительно меньше. Более того, очень часто в одной и той же кладке встречаются две (или даже три) порции икринок, резко различающихся по степени своего развития (например, одна порция икры может быть на стадии начала дробления,в то время как другая порция — на стадии начала гаструляции). Точно так же и среди личинок одного помета в ряде случаев без труда можно выделить две (а подчас и три) разновозрастные группы, различающиеся по длине тела на 7—8 мм. Обычно в подобных случаях различия в степени развития составляют 1—3 суток, а подчас и более. По-видимому, либо несколько самок последовательно откладывают свою икру в одну и ту жу выводковую камеру, либо одна и та же самка откладывает ее порционно через довольно значительные промежутки времени.
Выклюнувшиеся личинки с помощью цементной железы прикрепляются к стенкам выводковой камеры, где и висят почти неподвижно, пока у них не рассосется желточный мешок. Наличие четырех пар наружных жабр позволяет им обходиться без воздушного дыхания. Личинки растут очень быстро и уже через три недели достигают в длину 20—25 мм. К этому времени они утрачивают желточный мешок и переходят к активному питанию, поднимаясь к поверхности воды для дыхания атмосферным воздухом.
По достижении в длину 30—35 мм, немногим более чем через месяц после выклева, личинки навсегда покидают гнездо. К этому времени у них утрачивается одна пара наружных жабр. Остальные наружные жабры редуцируются очень поздно, и даже в течение нескольких лет у взрослых рыб сохраняются рудименты их базальных частей. До наступления засушливого периода личинки успевают достичь длину 70—120 мм, а способность зарываться в грунт для спячки и образовывать кокон они приобретают уже при длине тела 40—50 мм.
В неволе протоптеры очень нетребовательны и неприхотливы настолько, что могут жить в самой тухлой и мутной воде. Любопытно, однако, что в Нью-Йоркском аквариуме они не смогли жить в дехлорированной водопроводной воде. Лишь после того как эту воду продистиллировали, они почувствовали себя сносно. При соответствующем обращении протоптеры легко поддаются дрессировке. Так, например, если кормежку предварять стуком по стенке аквариума, то уже через 2—3 недели, заслышав сигнал, рыбы выказывают возбуждение и выходят к тому месту, где их ожидает пища. В противоположность миролюбивому американскому чешуйчатнику (Lepidosiren paradoxa) все виды протоптеров отличаются свирепым и неуживчивым нравом. Помещенные вместе, они не знают пощады и дерутся до тех пор, пока в живых не остается удачливый победитель. Если же к протоптеру подсадить каких-либо других крупных рыб, которых он заведомо не может использовать в пищу, то тем по менее он преследует их и калечит. Только молодых протоптеров, когда нет иного выхода, можно держать вместе. Но рано или поздно они так яростно нападают друг на друга, что вскоре оказываются без плавников. К счастью, откушенные плавники восстанавливаются у них очень быстро.
Обычно в аквариумы Европы и Америки протоптеров доставляют в коконе. Такой способ транспортировки чрезвычайно удобен, но требует большой осторожности, так как из-за встряски кокон легко может порваться, что приводит к неминуемой гибели рыбы. Замечательно также, что в тех случаях, когда кокон впавшей в спячку рыбы соприкасается не с грунтом, а с каким-либо инородным телом (например, со стеклянной стенкой аквариума), то это неизбежно приводит к смертельному исходу. Вот почему в искусственных условиям нижнюю часть стенки аквариума необходимо обмазывать толстым слоем глины.
Если потревожить протоптера в его “спальном гнезде”, то он издает звуки, напоминающие одновременно и писк и скрип, что, по всей видимости, связано со “скрежетом зубовным” в буквальном смысле этого слова. Раздраженная рыба вне воды способна издавать звуки, похожие на громкий вскрик. Такой же звук раздается тогда, когда из легких пойманной рыбы с силой выбрасывается воздух. В природных условиях при дыхании атмосферным воздухом протоптер издает громкий вздох, нередко переходящий в своеобразный визг, слышимый на далеком расстоянии.
Во многих районах Африки местное население промышляет протоптеров, так как мясо их отличается превосходными вкусовыми качествами. Наиболее легко добывать этих рыб во время спячки. Естественно, что для этого необходимо знать месга, где они залегают в спячку. Оказывается, жители Гамбии могут обнаруживать эти места на слух, так как, по их утверждению, в тихую погоiy на значительном расстоянии можно слышать, как дышит зарывшийся в землю крупный “кам''•опа” (P. annectens). Никому из исследователей ; этом отношении не посчастливилось.
Но свидетельству многих исследователей, оригинальный способ лова протоптеров применяется жителями Судана. Они пользуются специальным барабаном, с помощью которого извлекаются щуки, имитирующие падение дождевых капель. Поддавшись обману, протоптеры просыпаются ;i издают громкий чмокающий звук, выдавая гем самым место своего укрытия, а подчас даже и выползают из своих гнезд, попадая прямо в руки ловцов.
Американский чешуйчатник, или лепидосирен, Lt.'pidosiren paradoxa), населяет центральную часть Южной Америки. Его ареал охватывает почти весь бассейн Амазонки и северных притоков Параны. По строению и образу жизни лепидосирен очень похож на своих африканских сородичей. По сравнению с протоптерами его тело еще более вытянуто в еще большей степени напоминает тело угря, парные плавники еще более недоразвиты (в них полностью исчезают боковые хрящевые опорные элементы) и укорочены, чешуя еще глубже запрятана в кожу и еще мельче. Эта крупная рыба, достигающая в длину 125 см, окрашена в сероватобурые тона с большими черными пятнами на спине.
Образ жизни лепидосирена в основных чертах также очень схож с образом жизни протоптеров. Он, как правило, населяет лишь. временные заболоченные водоемы, сильно заросшие водной растительностью.Особенно он многочислен в таких водоемах, встречающихся во множестве на равнинах Гран-Чако. Эти водоемы наполняются водой в период тропических ливней (с апреля по сентябрь) и, как правило, высыхают в период засухи, приходящейся на остальную часть года.По мере усыхания водоема и по мере уменьшения в воде количества кислорода лепидосирен все чаще и чаще прибегает к дыханию атмосферным воздухом. Когда же слой воды становится совсем небольшим, он роет себе “спальное гнездо” и залегает в спячку, переходя полностью на дыхание атмосферным воздухом. По своей форме “спальное гнездо” лепидосирена ничем не отличается от “спального гнезда” протоптера и, как и последнее, состоит из расширенной “спальни” и воздушной (или входной) камеры, прикрытой сверху предохранительным колпачком. Помимо верхнего колпачка, в воздушной камере у лепидосирена иногда имеется дополнительная пробка из грунта. Изредка встречаются гнезда даже с двумя дополнительными пробками. Залегший в “спальню” лепидосирен принимает точно такое же положение, как и протоптер, однако в отличие от последнего он, по-видимому, не способен образовывать кокон. Правда, еще ни разу не удавалось обнаружить его гнездо в просохшем грунте: по крайней мере, на уровне “спальни” грунт всегда остается влажным, и, как правило, в ней сохраняется вода, смешанная со слизью, выделяемой спящим животным. В годы, обильные осадками, временные водоемы подчас не высыхают даже в период засухи и лепидосирен не впадает в спячку.
С началом периода дождей, когда пересохшие водоемы заполняются водой, лепидосирен покидает свое “спальное гнездо” (причем делает он это столь же осторожно и осмотрительно, как и протоптер) и набрасывается на пищу, проявляя необычайную прожорливость. Питается он различными беспозвоночными животными и главным образом крупными улитками ампуллариями. По-видимому, растительная пища играет немалую роль в его рационе, особенно у молоди. Почти все свое время лепидосирен проводит па дне, где он пли лежит неподвижно или медленно ползает на брюхе среди густых зарослей растительности. Время от времени он поднимается к поверхности для дыхания атмосферным воздухом. Сначала он высовывает рыло из воды и делает выдох. Затем на короткое время скрывается под водой и, снова выставив рыло, делает глубокий вдох. После этого животное медленно погружается на дно. выпуская избыток воздуха через жаберные отверстия.
Не проходит и двух-трех недель после окончания спячки, как лепидосирен уже приступает к размножению. Так же как и протоптер, к этому времени он роет выводковое гнездо, которое представляет собой довольно глубокую нору шириной 15—20 см с одним выходом, идущую обычно вертикально вниз и имеющую горизонтальное колено, которое заканчивается расширением. Обычно такие норы достигают в длину 60—80 см, но нередки случаи, когда они имеют в длину 1—1,5 м. Икринки диаметром 6,5 — 7,0 мм откладываются на отмершие листья и траву, которые специально затаскиваются в выводковую камеру. Охрану гнезда и потомства берет на себя самец. В период нереста у него на брюшных плавниках развиваются многочисленные ветвящиеся выросты длиной 5—8 см, пронизанные многочисленными кровеносными сосудами. Функциональное назначение этих образований еще не совсем ясно. По одной версии, через них из крови выделяется кислород и создаются более благоприятные условия для развития икры и личинок. По другой версии, наоборот, эти выросты играют роль дополнительных жабр, так как самец, охраняющий гнездо, не выходит на поверхность и лишен возможности дышать атмосферным воздухом. После того как самец покидает гнездо, эти выросты на брюшных плавниках редуцируются и сохраняются в виде небольших бугорков. Слизь, покрывающая тело чешуйчатника, обладает коагулирующими свойствами и способна очищать воду от мути. Это благотворно сказывается на развитии икры и личинок.
Личинки лепидосирена, так же как и личинки протоптеров, имеют наружные жабры и цементную железу, с помощью которой они подвешиваются в гнезде. Личинки растут довольно быстро: через два месяца после выклева, т. е. к моменту рассасывания желточного мешка и перехода на активное питание, они достигают в длину 55 мм. Однако дышать атмосферным воздухом личинки начинают еще задолго до этого (при длине 32—40 мм), когда они еще пребывают в гнезде под охраной самца. Наружные жабры исчезают у них вскоре после того, как они покидают гнездо. По окончании нереста лепидосирен продолжает усиленно отъедаться, восполняя потери, понесенные во время спячки и нереста, и создавая запасы жира на время предстоящей спячки. В отличие от протоптеров во время спячки он расходует жир, который откладывается впрок в больших количествах в межмышечных тканях. Имеются сведения, что эта рыба способна издавать звуки, напоминающие кошачье мяуканье.
Индейцы преследуют лепидосирена ради его вкусного мяса. В неволе лепидосирен очень неприхотлив, миролюбив и легко уживается с другими рыбами.